


我国变应性鼻炎的发病率呈逐年上升的趋势,鼻用糖皮质激素被认为是目前治疗变应性鼻炎的一线用药。由于鼻用糖皮质激素药物的广泛应用,临床上患者的激素治疗效果存在显著的个体差异,有些患者出现激素治疗无效的情况,这种情况我们称为糖皮质激素抵抗。
我们先来了解一下糖皮质激素的作用机制(图1),糖皮质激素(Glucocorticoid GC)是脂溶性类固醇激素,内源性的糖皮质激素由肾上腺皮质束状带分泌,受下丘脑-垂体-肾上腺轴调节。外源性或内源性的糖皮质激素通过细胞膜扩散进入细胞质中,与糖皮质激素受体(Glucocorticoid Receptor GR)结合转位进入细胞核,与相应靶基因的上游启动子的GRE(Glucocorticoid Responsive Element,糖皮质激素反应元件)结合,通过促进或抑制相关基因的转录而发挥抗炎及免疫抑制等作用。糖皮质激素受体(GR)几乎在所有组织和细胞中表达。上述糖皮质激素作用途径中任何过程出现问题均可影响糖皮质激素发挥作用而导致糖皮质激素抵抗。

图1 糖皮质激素作用机制
1.糖皮质激素受体前因素:
(1)皮质类固醇结合球蛋白(CBG)的变异:
皮质类固醇结合球蛋白(CBG)是内源性糖皮质激素(GC)血浆的载体蛋白,可以保持GC的失活状态。血浆内80%的糖皮质激素(GC)与皮质类固醇结合球蛋白(CBG)结合,有10-15%与血浆清蛋白松散结合,有5-8%处于游离状态,目前认为处于游离状态的GC具有生物学活性。在机体炎症反应时皮质类固醇结合球蛋白(CBG)与GC解离,释放GC,来调节靶组织中GC的量。如果皮质类固醇结合球蛋白(CBG)变异,就可以影响GC的效应。
(2)多药耐药基因(MDR-1):
多药耐药基因(MDR-1)编码蛋白p-糖蛋白170,这种蛋白表达于淋巴细胞和多种细胞表面,能主动把内源性或外源性糖皮质激素排出靶细胞,若p-糖蛋白170增多可导致GC不能进入细胞,从而降低GC的疗效。
2.糖皮质激素受体因素:
外源性或内源性的糖皮质激素通过细胞膜扩散进入细胞质中,与糖皮质激素受体(GR)结合转位进入细胞核,通过促进或抑制相关基因的转录而发挥抗炎及免疫抑制等作用。理解糖皮质激素与糖皮质激素受体的结合,首先需要掌握糖皮质激素受体(GR)结构(图2)。
糖皮质激素受体(GR)是核受体超家族成员之一,其编码基因位于染色体5q31,是一个配体(GC)依赖转录因子。糖皮质激素受体(GR)分子结构由与转录活化有关的N末端区、中心高度保守的含2个锌指的DNA结合区(DBD)及C端配体结合区(LBD)组成,配体就是糖皮质激素(GC)。
N末端区含有一个与靶基因转录活化有关的反式激活结构域AF21。
DNA结合区第一个锌指可与核因子κB(NF-κB)结合,第二个锌指可与GR二聚体化及与靶基因糖皮质激素反应元件(GRE)结合有关,DNA结合区还含有核定位信号(NLS1)。
C端配体结合区(LBD)除与糖皮质激素(GC)结合外,还与热休克蛋白(HSP)结合,并且含有第二个核定位信号(NLS2)和反式激活结构域AF22。糖皮质激素受体本身或与其相关的蛋白分子出现问题均会出现糖皮质激素抵抗。
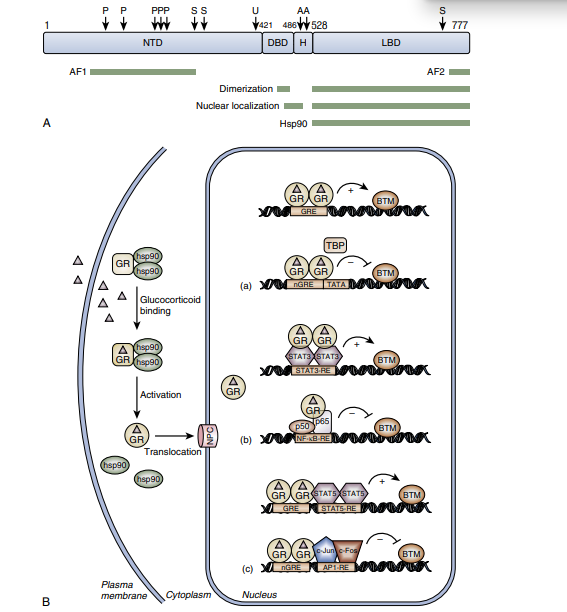
糖皮质激素受体(GR)基因经选择性剪接可生成5种亚型:GRα、GRβ、GRγ、GRρ、GR-A,它们有各自不同的生物学活性。GRα、GRβ是主要的两种受体,α型属于功能型,β型属于拮抗型。生理条件下以GRα为主要形式。GRα是经典的糖皮质激素配体结合蛋白,与糖皮质激素结合后调节细胞抗炎基因的表达,而GRβ由于缺乏完整的激素结合区,与激素结合形成无DNA转录能力的异源二聚体,与GRα竞争激素反应元件的DNA序列靶位,从而从转录水平上减弱糖皮质激素的效应。所以有些学者通过研究糖皮质激素抵抗模型中GRα和GRβ这两种受体的异常表达,GRα/GRβ比率的降低来寻求导致糖皮质激素抵抗的原因及干预机制。GRρ有激活GRα的作用;糖皮质激素抵抗的早期会诱导GR-A的产生,一旦到了稳定期,GR-A的表达将受到限制。GRγ、GRρ、GR-A三种亚型研究较少,具体生物学活性还不确定。目前认为糖皮质激素抵抗可能与糖皮质激素受体的表达量,各亚型之间的比例,基因突变,基因多态性等多种因素有关。
糖皮质激素受体(GR)在未与糖皮质激素(GC)结合时位于细胞质内,以大分子多蛋白复合物形式存在。此复合物由GR、两分子HSP90和另外几个受体相关蛋白组成。当糖皮质激素(GC)进入细胞内,该复合物通过磷酸化作用发生构型改变,糖皮质激素受体(GR)与分子伴侣HSP90解离,发生二聚化,暴露出DNA结合区,并与糖皮质激素(GC)结合。GR-GC复合物转位进入细胞核后,通过促进或抑制相关基因的转录而发挥抗炎和免疫抑制等作用。进一步研究发现,当热休克蛋白(HSP90)增多会降低糖皮质激素受体(GR)由细胞核回细胞质的速度,使糖皮质激素受体(GR)在核内滞留,导致糖皮质激素(GC)进入细胞核减少,产生糖皮质激素抵抗。
3.糖皮质激素受体后因素:
是细胞核内与糖皮质激素受体(GR)基因突变或者与其它调节糖皮质激素受体(GR)功能的基因多态性相关的因素。主要是位于不同染色体上的不同基因,如表1,具体的变异位点及影响糖皮质激素疗效的机制还未研究清楚。
表1 糖皮质激素受体后因素
|
基因名称 |
染色体位置 |
|
糖皮质激素受体基因 GR/NR3C1 |
5q31 |
|
哮喘候选基因TBX21 |
17q21.32 |
|
肾上腺皮质激素释放激素受体1基因 CRHR1 |
17q21.31 |
|
糖皮质激素诱导转录因子1基因 GLCCI1 |
7p21.3 |
|
免疫球蛋白E低亲和力片段Ⅱ基因 FcER2 |
19p13.3 |
与糖皮质激素作用相关的细胞信号通路(如下图):
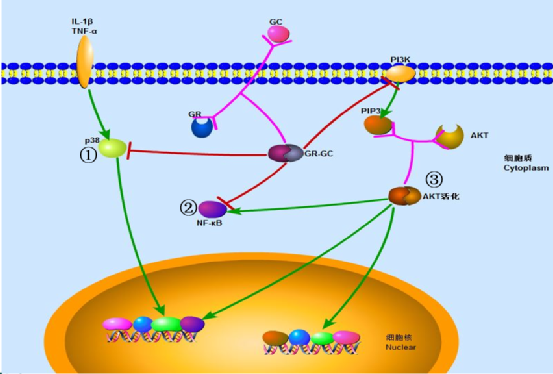
①MAPK p38-JNK信号转导通路(丝裂原活化蛋白激酶-c-Jun氨基末端激酶):前炎症因子IL-1β、TNF-α可激活丝裂原活化蛋白激酶p38,导致GR-GC复合物出细胞核转运,还可以进而诱导细胞核内下游细胞通路的转导及DNA的转录,导致炎症细胞分泌大量炎症因子。正常条件下,GR-GC和丝裂原活化蛋白激酶p38可以相互抑制,达到平衡,当糖皮质激素受体异常或大量前炎症因子刺激后,糖皮质激素效应降低,出现糖皮质激素抵抗。
②NF-κB信号通路:它是MAPK p38-JNK信号转导通路(丝裂原活化蛋白激酶-c-Jun氨基末端激酶)的下游通路之一,在核内NF-κB、AP1(活化蛋白1)和糖皮质激素受体(GR)相互竞争转录的共刺激分子CBP/p300,在细胞质内GR-GC复合物可以抑制NF-κB核转位入核。若各种因素导致糖皮质激素效应减弱时,会出现平衡打破,NF-κB核转位入核增多,导致炎症细胞分泌大量炎症因子,形成恶性循环。
③PI3K-AKT信号通路(磷脂酰肌醇3激酶-蛋白激酶B):是人体很多细胞发挥增殖、分化、凋亡和葡萄糖转运等多种功能调节机制的经典信号通路。磷脂酰肌醇3激酶(PI3K)在细胞膜上被激活产生第二信使PIP3(三磷酸磷脂酰肌醇),PIP3(三磷酸磷脂酰肌醇)可以与含有PH结构域的信号蛋白AKT结合,并活化AKT(蛋白激酶B)。AKT是抗细胞凋亡因子,活化的AKT又可以激活下游的许多转录因子,其中与炎症相关的有AP1(活化蛋白1)、CRE(cAMP反应元件)、NFAT(活化T细胞核因子)及GRE(糖皮质激素反应元件)。这些下游通路有些可以促炎,有些可以抗炎。由于目前没有公认的糖皮质激素抵抗模型,所有试验是在单一的H2O2刺激的体外单核细胞模型上完成,没有体内试验的支持,所以糖皮质激素在这条细胞通路上是发挥抑制作用,还是促进作用还有待继续研究。
1.糖皮质激素受体的减少;
2.GRα/GRβ比例失衡;
3.糖皮质激素基因突变;
4.HSP90(热休克蛋白)的异常;
5.GR-GC复合物入核异常;
6.T细胞异常导致炎症因子大量分泌,可以降低糖皮质激素受体(GR)与糖皮质激素(GC)亲和力。

